BioGeoChemistry of Tidal Flats
Identification of abundant and active sulfate-reducing bacteria in deep sediments
BACKGROUND
Previous studies on tidal surface sediments in the German Wadden Sea have identified sulfate-reducing bacteria (SRB) to be highly abundant and active in near-surface layers where microbial activity is not limited by the availability of easily degradable electron donors (Llobet-Brossa et al., 1998, 2000; Mussmann et al., 2005). In contrast, deeper sediment layers are assumed to be less active due to the increasing amount of recalcitrant organic matter and decreasing cell numbers.
However, the dynamical structure of sandy tidal flats (e.g. site 'Janssand') facilitates advective pore water transport and may therefore lead to the provision of nutrients into greater depths (Billerbeck et al., 2006). Thus, this project addressed the question whether this dynamic feature is reflected by the abundance, activity and community structure of SRB.
Selective cultivation conditions were chosen to facilitate growth of abundant, either lactate- acetate- or hydrogen-utilizing SRB from several meters deep sediments. Their cultivation and final isolation was accompanied and evaluated by molecular techniques (DGGE, cloning and sequencing). Furthermore, the abundance of cultured SRB was verified by a cultivation-independent in situ hybridization technique (CARD-FISH, catalyzed reporter deposition fluorescence in situ hybridization). Enumeration was performed with a set of six oligonucleotide CARD-FISH probes targeting widely distributed groups of deltaproteobacterial SRB as well as relatives to Desulfotomaculum spp., a spore-forming firmicute. As these probes target (transcribed) 16S rRNA, it is assumed that only "active" cells are detected with a detection limit of ~50 targets (ribosomes) per cell.
RESULTS
In contrast to previously studied mud flat sediments, total cell counts varied only slightly with depth (~3-6.5 x 108 cells cm-3 sediment, Fig. 1B). Bacteria dominated the microbial community accounting for 60-98% of the total cell counts. With respect to the low concentrations of pore water sulfate (<0.1 mM from 1.5 meters downwards), SRB detected with the chosen set of probes were unexpectedly numerous down to 5 meters depth. Their relative abundance even increased from ~2.5% in the sediment surface to ~7% in 5 meters depth (Fig.2A).
Consistently, both cultivation and CARD-FISH analysis showed that certain members of the Desulfobacteraceae and the Desulfobulbaceae predominated the sulfate-reducing community either related to Desulfobacter sp. and Desulfobacula sp. (Desulfobacteraceae) or Desulfobacterium corrodens and D. catecholicum (Desulfobulbaceae). They were not only isolated from up to million-fold diluted cultures, but appeared to be highly abundant in situ. At least two-third of all SRB in sediments below 50 cm were targeted by the corresponding CARD-FISH probes (Sval, 428, DSB98) what was particularly true for the depths sampled for cultivation (Fig. 2B).
As strains affiliated with Desulfobacterium corrodens were most frequently isolated from different depths and with various substrates, they were chosen for more detailed phylogenetic and physiological characterization. These strains shared at least 98 % 16S rRNA gene sequence identity with each other and 95-96.6 % identity with the not validly described 'Desulfobacterium corrodens' strain IS4. Desulfopila aestuarii was identified as the closest cultured and validly described related organism (< 93 %) meaning that our isolates belong to another, and apparently phylogenetically novel, genus of deltaproteobacterial SRB. They were shown to utilize a variety of organic compounds (e.g. short-chain fatty acids, alcohols) and molecular hydrogen as electron donor for sulfate reduction. Their capability to oxidize more complex or 'unusual' substrates (e.g. aromatic compounds, Fe(II)) is currently under investigation.
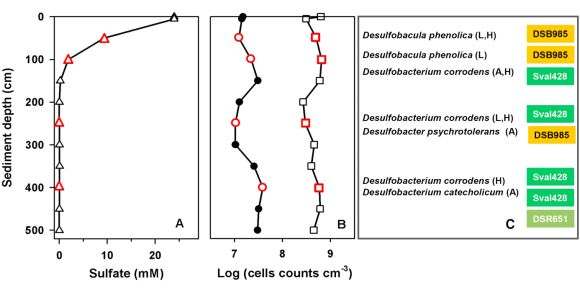
Fig.1 Depth profiles of (A) sulfate from pore water samples, (B) total cell counts (DAPI; open squares) and absolute abundance of SRB as detected with the chosen set of six CARD-FISH probe (filled circles). Red symbols indicate sediment depths that were sampled for cultivation of abundant SRB. (C) Closest cultured relatives to the isolated SRB and corresponding CARD-FISH probes. Substrates used for isolation are given in brackets (L = lactate, A = acetate, H = hydrogen).
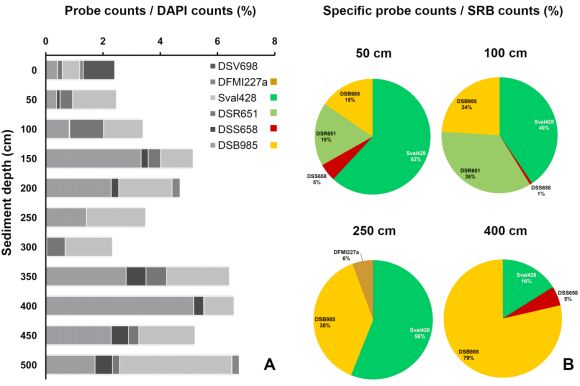
Fig.2 (A) Relative abundance of different phylogenetic groups of SRB to the total microbial community.
(B) Relative abundance of different phylogenetic groups of SRB to the total SRB community in sediment depths that were sampled for cultivation of abundant SRB.
(Probes and targets: Sval428, DSR651: Desulfobulbaceae; DSS658, DSB985: Desulfobacteraceae; DSV986: Desulfovibrio spp.; DFMI227a: Desulfotomaculum spp.)
CONCLUSION
Extending the knowledge about the abundance and vertical distribution of SRB in tidal sand flats, our results support the hypothesis that deep sediments have a so far unrecognized impact on biogeochemical cycling within the tidal flat ecosystem. The relatively high abundance of microbial cells and the presence of active and abundant SRB might generally be regarded as indication for a beneficial effect of pore water drainage and the provision of substrates to the anaerobic community in several meters deep sediments. SRB might additionally overcome sulfate depletion, that was observed over seasons, by energy conservation via alternative metabolic pathways, e.g. mutualistic associations with other anaerobes.
People involved in this project:
Antje Gittel, Stephanie Reischke, Martin Koenneke, Heribert Cypionka
Henrik Sass (University of Cardiff), Marc Mussmann (MPI Bremen)
| <<back | All publications
|